La adquisición mitocondrial y los inicios de la célula eucariota: dos perspectivas temporales
Daniela Orozco Pérez, Adrián Díaz-Chávez, Christian Antonio Basurto Muñoz y Rafael Montiel****
El proceso por el cual surgieron las células eucariotas, llamado eucariogénesis, es uno de los mayores eventos evolutivos en la historia de la vida; uno de los más enigmáticos del campo biológico. Un momento crucial en este proceso fue la aparición de las mitocondrias, organelos conocidos como “los motores de la célula”, ya que son sumamente relevantes para el funcionamiento de la célula eucarionte. Se encargan de la generación de la energía celular (en forma de moléculas de “Adenosín Trifosfato” o ATP), así como de muchos otros procesos vitales. Son portadoras del ADN mitocondrial, que codifica genes esenciales para la producción de ATP y las alteraciones en su maquinaria pueden contribuir al desarrollo de enfermedades metabólicas, neurodegenerativas y cáncer, así como al proceso de envejecimiento. El conocimiento del origen de la mitocondria y su papel en el surgimiento de la célula eucariota nos permitirá comprender mejor su biología, diseñar herramientas de diagnóstico más precisas, e incluso acercarnos a la creación de vida artificial, con gran potencial biotecnológico.
Los trabajos pioneros de Wallin y Andreas Schimper explican el origen bacteriano de la mitocondria, y de otros organelos, a través de eventos de endocitosis[1] (Wallin, 1925). Esta idea fue desarrollada de forma independiente por Lynn Margulis en 1967, quien propuso que la mitocondria, así como otros organelos, provenían de procariontes. Planteó también que estos organelos habrían sido generados por eventos de engullimiento entre dos especies bacterianas y que esto sería un motor de la evolución, contribuyendo a la generación de nuevas especies (Sagan, 1967). La interacción íntima entre organismos de diferentes especies es conocida como simbiosis e implica contacto físico en una relación prolongada. La teoría endosimbiótica plantea que esta relación se dio de tal forma que uno de los organismos existía dentro de otro hasta convertirse en organelo.
La endosimbiosis que daría lugar al origen de la mitocondria se considera un evento evolutivo único (Zachar, 2020), pero ¿cómo se llevó a cabo tal adquisición?, y aún más intrigante, ¿cuándo llegó el endosimbionte protomitocondrial? Al respecto, se han conformado dos hipótesis generales, que coinciden en el origen bacteriano de la mitocondria, pero difieren en el tiempo de ocurrencia y la naturaleza del organismo receptor o anfitrión: las hipótesis mito-temprana y mito-tardía. La primera considera que la mitocondria se incorporó tempranamente en el proceso de eucariogénesis, por lo que debió de integrarse en un anfitrión simple. En contraste, la segunda argumenta que existió una complejidad mayor del anfitrión antes del arribo de la mitocondria, es decir, que se encontraba en un estadio avanzado de la eucariogénesis. Ambas hipótesis están embebidas en otras, como pueden ser la sintrófica o fagotrófica, que se refieren a cómo fue la interacción que permitió la adquisición mitocondrial, ya sea colaborativa o fagocitaria, respectivamente.
Aquí se describen los trabajos más relevantes acerca del origen de la mitocondria y su papel en la eucariogénesis, así como las perspectivas en torno al debate generado durante los últimos años sobre si la mitocondria fue adquirida tempranamente y fue esencial en el origen de la célula eucariota o si por el contrario fue una adquisición tardía, por una célula que ya estaba en un punto avanzado de la eucariogénesis.
La ventana de tiempo
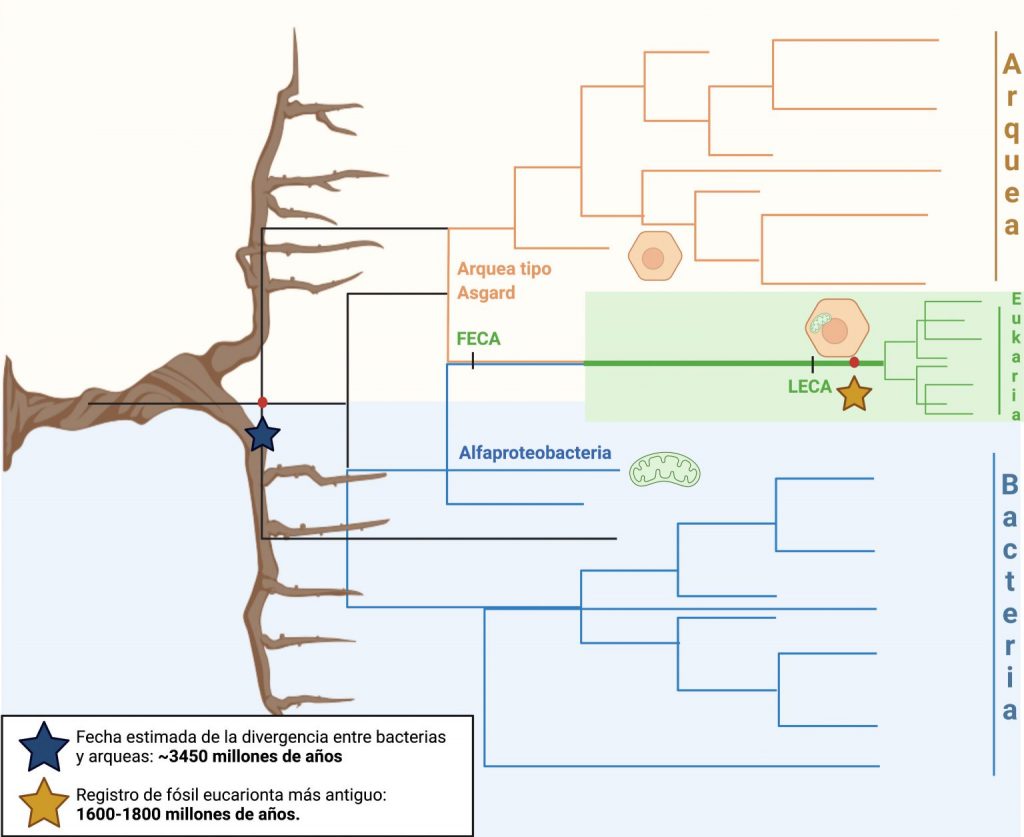
Para situarnos en el tiempo en que ocurrió la eucariogénesis y el surgimiento de la mitocondria, debemos preguntarnos primero, ¿cuántos dominios de la vida existen? La hipótesis de los tres dominios divide al árbol de la vida en “Bacteria”, “Archaea” y “Eukarya” como un solo grupo con un único ancestro definible (Woese et al., 1990). Por otro lado, la hipótesis de dos dominios primarios sugiere que en realidad los eucariontes forman una rama dentro de las arqueas (Koumandou et al., 2013). El descubrimiento de un nuevo linaje arqueal (Lokiarchaeota[2]), que representa al pariente más cercano de los eucariontes (Spang et al., 2015) y la descripción del superfilo Asgard (Zaremba-Niedzwiedzka et al., 2017), apoyan la hipótesis de los dos dominios. No obstante, la metodología usada en la descripción de los Asgard ha sido criticada (Da Cunha et al., 2018) dando entonces apoyo al modelo de tres dominios primarios, por lo que la relación entre Archaea y Eukarya sigue siendo una pregunta abierta en espera de resolverse. Lo que sí está claro es que Eukarya es un grupo diferenciado con características específicas que comparten todos sus miembros, así como las arqueas también pueden diferenciarse de las bacterias.
También es importante definir al primer ancestro común de los eucariotas (FECA, por sus siglas en inglés), que representa al organismo más antiguo cuyos únicos descendientes vivos son los eucariotas; así como a su último ancestro común (LECA, por sus siglas en inglés), que equivale a su antecesor más reciente (Eme et al., 2017). La estimación de las fechas en las que existieron estos ancestros depende de varios factores. Por ejemplo, el fósil eucariota más antiguo está datado en alrededor de 1600-1800 millones de años (Pang et al. 2013), por lo que LECA no podría ser más reciente que esa fecha, pero sí más antiguo, lo que sería evidenciado por el descubrimiento de fósiles eucariotas más lejanos. Por otro lado, la antigüedad máxima de FECA está limitada por la separación evolutiva que ocurrió entre las arqueas y las bacterias, estimada actualmente en 3450 millones. Entonces, con los datos que se tienen actualmente, la ventana temporal donde se podrían situar FECA y LECA, y en la cual se daría la adquisición del simbionte protomitocondrial, estaría delimitada entre los 3500 y los 1600 millones de años (Figura 1).
La hipótesis mito-temprana
El modelo clásico de la hipótesis mito-temprana es la hipótesis del hidrógeno (Figura 2), propuesta por Martin y Muller (1998). Postula que una arquea anaeróbica asimiló a una alfa-proteobacteria capaz de respirar, pero que generaba hidrógeno molecular como desecho y de la cual dependía su metabolismo. En esta propuesta no se requiere que el anfitrión poseyera alguna estructura compleja como núcleo o citoesqueleto.
Con este antecedente, Moreira y López-Garcia (1998) proponen un modelo en el que participan un arqueón metanogénico y una myxobacteria (una delta-proteobacteria) para configurar un consorcio metabólico en el que se incorporaría tempranamente la alfa-proteobacteria que resultaría en la mitocondria (Figura 2). Plantean una myxobacteria, porque presenta diversas similitudes con eucariontes en los niveles molecular y metabólico y en el complejo ciclo de desarrollo y división celular. Sin embargo, una de las debilidades de esta hipótesis es que los métodos filogenéticos no muestran una evidencia sólida de que una delta-proteobacteria haya sido ancestro de las eucariotas, aunque la antigüedad de esta relación pudo borrar la señal filogenética.
Las observaciones obtenidas con métodos metagenómicos también favorecen una adquisición mito-temprana. Martijn y col. (2018) analizaron una mayor diversidad de organismos oceánicos, que no eran cultivables en el laboratorio, y lograron reconstruir 45 genomas que representan 12 nuevos linajes alfa-proteobacterianos. Su posición filogenética sugirió que la mitocondria tiene en realidad un origen más antiguo que el pensado hasta entonces, anterior, aunque cercano, a la diversificación de las alfa-proteobacterias. Este resultado apoya una adquisición mito-temprana.
Entre los trabajos más recientes, la propuesta hecha por López-García y Moreira (2020) toma en cuenta los nuevos descubrimientos de arqueas del grupo Asgard; un linaje de ramificación antigua que comparte más genes similares a eucariotas que otras arqueas. El modelo propone una endosimbiosis tripartita del grupo Asgard con una delta-proteobacteria reductora de sulfato, formando un protoeucarionte que en seguida asimila una alfa-proteobacteria aeróbica facultativa y oxidadora de sulfatos, de la cual se deriva la mitocondria, y en conjunto inician la eucariogénesis.
Algunos grupos de investigación usan microfósiles y su datación geológica para calibrar la escala temporal del árbol de la vida. Por ejemplo, Betts y col. (2018), emplearon microfósiles de diferentes eras geológicas para precisar la temporalidad en la que los distintos organismos, desde LUCA y cianobacterias, hasta eucariotas, aparecieron en la Tierra. Los resultados acerca de la eucariogénesis y la adquisición mitocondrial suponen una temporalidad similar (alrededor de 1,679-1,866 millones de años) tanto para las alfa-proteobacterias como para LECA, apoyando así la hipótesis mito-temprana.
La hipótesis mito-tardía
A pesar del impacto de los múltiples trabajos que apoyan un escenario mito-temprano, recién han surgido contrapartes contundentes que sacuden los cimientos de las teorías planteadas anteriormente.
En el escenario mito-tardío la mitocondria no interviene en el inicio de la eucariogénesis, pero sí se propone como un elemento fundamental para el éxito evolutivo de las eucariotas. En efecto, Pittis y Gabaldón (2016) postulan que la endosimbiosis mitocondrial fue uno de los últimos pasos en la eucariogénesis, pero que este hecho proveyó de una ventaja adaptativa definitiva sobre aquellas formas menos complejas que carecían de mitocondria. Los experimentos que los llevaron a estas conclusiones se basan en árboles filogenéticos que realizaron con grupos de proteínas mitocondriales y no mitocondriales, comparándolos con proteínas de los otros dos dominios de la vida (Archaea y Bacteria). Encuentran que las proteínas mitocondriales presentan los árboles con los tallos más cortos, en comparación con las proteínas nucleares, lo que indicaría que las primeras se adquirieron más recientemente que las segundas si se asume que la longitud de los tallos refleja la distancia evolutiva. Por lo tanto, sus observaciones apoyan la adquisición tardía de la mitocondria, mientras que otras características eucariotas importantes se habrían obtenido anteriormente.
Esta visión es compatible con la idea de que un anfitrión complejo protoeucarionte o un anfitrión complejo arqueal, ya albergaba vías metabólicas complejas de origen bacteriano (Figura 2). De hecho, las reconstrucciones en el ámbito molecular de los principales sistemas celulares predicen que LECA poseyó una configuración de complejidad remarcable (Koumandou et al., 2013). Pero ¿por qué LECA tendría que ser tan complejo? La respuesta más sencilla es que se requería cierto nivel de complejidad para dominar el paisaje eucariótico temprano y ocupar una posición exitosa dentro del ecosistema (Koumandou et al., 2013) y además así tendría plena capacidad fagocitaria (Zachar et al., 2020).
En la corriente mito-tardía se identifican otros trabajos relevantes. Stairs (2020) encontró un organismo adicional como colaborador importante durante la evolución eucariota: un grupo anaeróbico conocido como anoxiclamidae al que identificó como el pariente más cercano de elementos del metabolismo del hidrógeno en eucariontes (Figura 2). Esto apoyó la hipótesis tardía de la mitocondria y su naturaleza mosaica, producto de la transferencia horizontal de genes. Por otra parte, y considerando que el origen de la mayoría del proteoma mitocondrial no proviene de las alfa-proteobacterias, Gray (2014) plantea que el anfitrión ya contaba con un organelo (premitocondria) de origen endógeno, que aportaría la mayoría de los componentes no alfa-proteobacterianos relevantes, aunque no todos los elementos necesarios para la formación de la mitocondria actual (Figura 2). Este organelo endógeno tendría un papel metabólico relacionado con vías que ahora son propias de la mitocondria, como el metabolismo de aminoácidos, nucleótidos y lípidos. Sin embargo, era un gran consumidor de energía, por lo que contaba con las vías necesarias para importar ATP, características que harían más fácil la asimilación del ancestro mitocondrial, propiciando subsecuentes rearreglos de elementos funcionales y estructurales para convertirse en el principal generador de energía celular; un organelo capaz de producir y exportar ATP. Esta hipótesis también es congruente con una adquisición mito-tardía.
Pese a estos resultados e hipótesis en favor de una adquisición tardía, hubo una pronta respuesta por parte de otros grupos de estudio, que no tardaron en mostrar su inconformidad con estos resultados, lo que avivó el debate. Martín y col. (2017) hicieron una dura crítica de la metodología y los supuestos que se utilizaron en el estudio de Pittis y Gabaldón (2016). A través de simulaciones probaron que se generaron errores por utilizar modelos que requieren estimar un número excesivo de parámetros, lo que se conoce como sobreparametrización. Argumentaron también que no es acertado suponer que la longitud del tallo de los árboles filogenéticos refleja la distancia evolutiva, desvalorizando dicho trabajo.
El anfitrión
Las controversias referentes a cómo se llevó a cabo la adquisición mitocondrial surgen de las incógnitas acerca del anfitrión, por lo que ha crecido el interés por caracterizar su naturaleza: ¿cómo era el organismo que integró eficazmente a un endosimbionte bacteriano?
Al aislar y secuenciar una arquea perteneciente al filo Asgard (Candidatus Prometheoarchaeum syntrophicum), Imachi y col. (2020) proponen un nuevo modelo de eucariogénesis: enredar-engullir-endogenizar (modelo E3). El modelo establece que la arquea era capaz de producir y transferir H2 y que interactuaba con una bacteria reductora de sulfatos. Esta interacción facilitaría la captura de una bacteria mediante estructuras externas que después favorecerían su engullimiento, formando una simbiosis primitiva que la transformaría en la mitocondria. Al ser aeróbica facultativa, esta bacteria aportaría al conjunto la capacidad de sobrevivir en presencia de oxígeno. Es el genoma arqueal más cercano a los eucariotas que se tiene a la fecha y es una de las pistas más recientes respecto a la identidad del anfitrión, aunque los mismos autores consideran que este modelo no es definitivo.
Por otra parte, Zachar y Boza (2020) no consideran posible una endosimbiosis procariota-procariota por la baja complejidad de los organismos participantes. Argumentan que no existen registros de que la simbiosis procariota-procariota tenga el mismo éxito que cuando participa un eucariota, y sugieren que no es posible que el único evento de endosimbiosis procariota-procariota conocido sea el caso de la mitocondria. Tampoco hay ejemplos contemporáneos de un anfitrión arqueal y un endosimbionte bacteriano, y aunque se conocen simbiosis muy cercanas entre bacterias (consorcios bacterianos), todos los eventos auténticamente endosimbióticos conocidos se dan con anfitriones eucarióticos. Los autores concluyen entonces que las mitocondrias debieron surgir de una interacción parasitaria o fagotrófica en la que participó un anfitrión más complejo, es decir, un organismo más cercano a una célula eucariota.
Apoyando la idea de que el anfitrión debió ser más complejo, Martijn y col. (2013) exponen la hipótesis de la arquea fagocitaria (PhAT por sus siglas en inglés), que está en acuerdo con la adquisición mito-tardía, ya que participa un protoeucarionte con un citoesqueleto suficientemente complejo y con la capacidad de fagocitar. Empero, se ha observado que Candidatus Prometheoarchaeum syntrophicum tiene un tamaño muy pequeño y carece de maquinaria y energía suficientes (Imachi et al., 2020), lo que hace poco probable que pudiera fagocitar, descartando esta hipótesis mito-tardía y apoyando en su lugar al modelo alternativo E3 descrito anteriormente. Como vemos, la simplicidad o complejidad del anfitrión y su significado para las hipótesis mito-tempranas o mito-tardías, es también un área actual de debate e investigación.
Conclusiones y perspectivas en un debate abierto
La información obtenida de los estudios ambientales y de biología celular, así como los recientes avances en herramientas filogenéticas y metagenómicas, permiten estar, cada vez más cerca, de reconstruir un modelo de eucariogénesis que represente de alguna manera este evento evolutivo. Una de las propuestas más recientes, dada por Bateman (2020), explora el modelo del “tercer espacio” (“third-space model”), en el cual expone que una matriz extracelular tipo biopelícula sirvió como andamio para poblaciones mezcladas de arqueas y bacterias que culminaría en la formación de una célula precursora.
Una aproximación interesante, que ha surgido como alternativa para estudiar la evolución de la eucariogénesis, es la biología sintética. En este campo, junto con organismos modificados, se intenta reproducir las condiciones en las que se originaron las células eucariotas. Se han recapitulado experimentalmente las primeras etapas de la evolución mitocondrial, a través de la generación de un endosimbionte (bacteria Escherichia coli) que le proporciona ATP a una célula de levadura deficiente de ésta (Mehta et al., 2018). Aunque estos experimentos no resuelven las incógnitas planteadas, nos conceden una pequeña ventana a la cual asomarnos en busca de pistas relativas al proceso de la eucariogénesis.
Efectivamente, la eucariogénesis y la llegada de la mitocondria ocurrieron y resultaron en una nueva complejidad representada por los eucariontes. Fue un evento evolutivo que trajo consecuencias que perduran hasta nuestros días, y, aunque queda mucho por investigar y un cúmulo de preguntas por responder, vivimos en una época emocionante por los avances tecnológicos y sus implicaciones para acercarnos a responder muchas de estas interrogantes.
Agradecimientos
Agradecemos a Luis José Delaye Arredondo (Unidad Irapuato, Cinvestav) por su asesoría experta, así como a Laila Pamela Partida Martínez (Unidad Irapuato, Cinvestav), Sean M. Rovito y Alexander de Luna Fors (ambos de la Unidad de Genómica Avanzada, Cinvestav) por sus valiosos comentarios. Este trabajo fue desarrollado originalmente durante el Curso de Razonamiento Científico Multidisciplinario (CRCM) del Programa de Posgrado en Biología Integrativa del Cinvestav en Irapuato.
Glosario
Adecuación biológica: Capacidad de un organismo, población o especie, de sobrevivir y reproducirse en el lugar donde se encuentra (Orr, 2009).
Alfa-proteobacteria: Perteneciente al grupo de organismos del filo proteobacteria; son diversos y suelen adoptar ambientes intracelulares (Williams et al., 2007).
Biopelícula: Poblaciones bacterianas incluidas en una matriz que se adhieren entre sí o a superficies o interfases (Costerton et al. 1995). Constituyen un ecosistema bacteriano.
Delta-proteobacteria: Subgrupo diverso de microorganismos gram-negativos perteneciente al filo proteobacteria. En el grupo hay géneros caracterizados por depredar otras bacterias (Madigan & Martinko, 1997).
Endocitosis: La captación de material extracelular en vesículas formadas a partir de la membrana plasmática (Cooper, 2000).
Eucariogénesis: Conjunto de eventos evolutivos que explican el proceso de aparición de las células eucariontes a partir de las células procariontes (Eme et al., 2017).
Fagocitosis: Captación celular de partículas (> 0,5 μ) dentro de una envoltura de membrana plasmática para su digestión (Hallett, 2020).
Hipótesis fagotrófica: Postula que una célula “protoeucarionte” (ni del tipo arquea, ni bacteria), contaba con mecanismos complejos propios de eucariontes como la capacidad de fagocitar, que se adquirieron y evolucionaron con anterioridad a la adquisición del ancestro mitocondrial, el cual fue engullido posteriormente por fagocitosis (Mills, 2020).
Hipótesis sintrófica: Se propone que los eucariotas evolucionaron a partir de una simbiosis metabólica tripartita basada en la transferencia de hidrógeno desde una delta-proteobacteria fermentativa a una arquea metanogénica y el reciclamiento de metano por parte de una alfa-proteobacteria aeróbica facultativa (López-García y Moreira, 2020).
Metagenómica: Un metagenoma representa una captura de pantalla genética de una comunidad, en un cierto tiempo y lugar, cuando se obtiene todo el ADN de ella y se analiza. Aproximación que requiere un buen diseño experimental y la combinación de diferentes herramientas tales como biología molecular, genética, bioinformática y estadística (Escobar-Zepeda et al., 2015).
Bibliografía
Bateman, A. (2020). Division of labour in a matrix, rather than phagocytosis or endosymbiosis, as a route for the origin of eukaryotic cells. Biology Direct, 15(1), 8. https://doi.org/10.1186/s13062-020-00260-9
Betts, H. C., Puttick, M. N., Clark, J. W., Williams, T. A., Donoghue, P. C. J., y Pisani, D. (2018). Integrated genomic and fossil evidence illuminates life’s early evolution and eukaryote origin. Nature Ecology and Evolution, 2(10), 1556–1562. https://doi.org/10.1038/s41559-018-0644-x
Cooper, G. M. (2000). The Cell: A Molecular Approach. 2nd edition. Sunderland (MA): Sinauer Associates. https://www.ncbi.nlm.nih.gov/books/NBK9926/
Costerton, J. W., Lewandowski, Z., Caldwell, D. E., Korber, D. R., y Lappin-Scott, H. M. (1995). Microbial biofilms. Annual Review of Microbiology, 49, 711-745. https://doi.org/10.1146/annurev.mi.49.100195.003431
Da Cunha, V., Gaia, M., Nasir, A., y Forterre, P. (2018). Asgard archaea do not close the debate about the universal tree of life topology. PLOS Genetics, 14(3), e1007215. https://doi.org/10.1371/journal.pgen.1007215
Eme, L., Spang, A., Lombard, J., Stairs, C., y Ettema, T. (2017). Archaea and the origin of eukaryotes. Nature Reviews Microbiology, 15(12), 711-723. https://doi.org/10.1038/nrmicro.2017.133
Escobar-Zepeda, A., Vera-Ponce de León, A., y Sanchez-Flores, A. (2015). The Road to Metagenomics: From Microbiology to DNA Sequencing Technologies and Bioinformatics. Frontiers In Genetics, 6, 348. https://doi.org/10.3389/fgene.2015.00348
Gray, M. W. (2014). The pre-endosymbiont hypothesis: A new perspective on the origin and evolution of mitochondria. Cold Spring Harbor Perspectives in Biology, 6(3), a016097. https://doi.org/10.1101/cshperspect.a016097
Hallett, M. B. (2020) An Introduction to Phagocytosis. Advances in Experimental Medicine and Biology, 1246, 1-7. https://doi.org/10.1007/978-3-030-40406-2_1
Imachi, H., Nobu, M., Nakahara, N., Morono, Y., Ogawara, M., Takaki, Y. et al. (2020). Isolation of an archaeon at the prokaryote–eukaryote interface. Nature, 577(7791), 519-525. https://doi.org/10.1038/s41586-019-1916-6
Koumandou, V., Wickstead, B., Ginger, M., van der Giezen, M., Dacks, J., y Field, M. (2013). Molecular paleontology and complexity in the last eukaryotic common ancestor. Critical Reviews In Biochemistry And Molecular Biology, 48(4), 373-396. https://doi.org/10.3109/10409238.2013.821444
López-García, P., y Moreira, D. (2020). The Syntrophy hypothesis for the origin of eukaryotes revisited. Nature Microbiology, 5(5), 655-667. https://doi.org/10.1038/s41564-020-0710-4
Madigan, M.T., Martinko, J.M. y Parker, J. (1997). Brock Biology of Microorganisms. 8th Edition. New York, USA: Prentice Hall International, Inc.
Martijn, J., y Ettema, T. J. G. (2013). From archaeon to eukaryote: The evolutionary dark ages of the eukaryotic cell. Biochemical Society Transactions, 41(1), 451-457. https://doi.org/10.1042/BST20120292
Martijn, J., Vosseberg, J., Guy, L., Offre, P., y Ettema, T. J. G. (2018). Deep mitochondrial origin outside the sampled alphaproteobacteria. Nature, 557(7703), 101–105. https://doi.org/10.1038/s41586-018-0059-5
Martin, W. F., Roettger, M., Ku, C., Garg, S. G., Nelson-Sathi, S., y Landan, G. (2017). Late mitochondrial origin is an artifact. Genome Biology and Evolution, 9(2), 373-379. https://doi.org/10.1093/gbe/evx027
Martin, W., y Müller, M. (1998). The hydrogen hypothesis for the first eukaryote. Nature, 392(6671), 37-41. https://doi.org/10.1038/32096
Mills, D. B. (2020). The origin of phagocytosis in Earth history. Interface Focus, 10(4), 20200019. https://doi.org/10.1098/rsfs.2020.0019
Mehta, A. P., Supekova, L., Chen, J. H., Pestonjamasp, K., Webster, P., Ko, Y., Henderson, S. C., McDermott, G., Supek, F., y Schultz, P. G. (2018). Engineering yeast endosymbionts as a step toward the evolution of mitochondria. Proceedings of the National Academy of Sciences of the United States of America, 115(46), 11796–11801. https://doi.org/10.1073/pnas.1813143115
Moreira, D., & López-García, P. (1998). Symbiosis Between Methanogenic Archaea and δ-Proteobacteria as the Origin of Eukaryotes: The Syntrophic Hypothesis. Journal of Molecular Evolution, 47(5), 517-530. https://doi.org/10.1007/pl00006408
Orr, H. A. (2009). Fitness and its role in evolutionary genetics. Nature Reviews Genetics, 10(8), 531–539. https://doi.org/10.1038/nrg2603
Pang, K., Tang, Q., Schiffbauer, J. D., Yao, J., Yuan, X., Wan, B., Chen, L., Ou, Z., y Xiao, S. (2013). The nature and origin of nucleus-like intracellular inclusions in Paleoproterozoic eukaryote microfossils. Geobiology, 11(6), 499-510. https://doi.org/10.1111/gbi.12053.
Pittis, A., y Gabaldón, T. (2016). Late acquisition of mitochondria by a host with chimaeric prokaryotic ancestry. Nature, 531(7592), 101-104. https://doi.org/10.1038/nature16941
Sagan, L. (1967). On the origin of mitosing cells. Journal of Theoretical Biology, 14(3), 225-274. https://doi.org/10.1016/0022-5193(67)90079-3
Spang, A., Saw, J., Jørgensen, S., Zaremba-Niedzwiedzka, K., Martijn, J., Lind, A. E., et al. (2015). Complex archaea that bridge the gap between prokaryotes and eukaryotes. Nature, 521(7551), 173-179. https://doi.org/10.1038/nature14447
Stairs, C. W., Dharamshi, J. E., Tamarit, D., Eme, L., Jørgensen, S. L., Spang, A., y Ettema, T. J. G. (2020). Chlamydial contribution to anaerobic metabolism during eukaryotic evolution. Science Advances, 6(35), eabb7258. https://doi.org/10.1126/sciadv.abb7258
Wallin, I. E. (1925). On the nature of mitochondria. IX. Demonstration of the bacterial nature of mitochondria. American Journal of Anatomy, 36(1), 131-149. https://doi.org/10.1002/aja.1000360106
Williams, K. P., Sobral, B. W., y Dickerman, A. W. (2007). A robust species tree for the Alphaproteobacteria. Journal of Bacteriology, 189(13), 4578-4586. https://doi.org/10.1128/JB.00269-07
Woese, C., Kandler, O., y Wheelis, M. (1990). Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya. Proceedings of the National Academy of Sciences, 87(12), 4576-4579. https://doi.org/10.1073/pnas.87.12.4576
Zachar, I., y Boza, G. (2020). Endosymbiosis before eukaryotes: mitochondrial establishment in protoeukaryotes. Cellular and Molecular Life Sciences, 77(18), 3503-3523. https://doi.org/10.1007/s00018-020-03462-6
Zaremba-Niedzwiedzka, K., Caceres, E., Saw, J., Bäckström, D., Juzokaite, L., Vancaester, E., et al. (2017). Asgard archaea illuminate the origin of eukaryotic cellular complexity. Nature, 541(7637), 353-358. https://doi.org/10.1038/nature21031
Referencias
[1] Con el propósito de ayudar al lector a familiarizarse con los conceptos clave de este artículo, las palabras en negrita se definen en el glosario
[2] Para información respecto a cómo se llevó a cabo este descubrimiento revisar el Recuadro.
*Licenciada en Ciencias Genómicas y estudiante de maestría en el Programa de Posgrado en Biología Integrativa del Cinvestav en Irapuato.
**Ingeniero en Biotecnología y estudiante de maestría en el Programa de Posgrado en Biología Integrativa del Cinvestav en Irapuato.
***Licenciado en Arqueología y estudiante de maestría en el Programa de Posgrado en Biología Integrativa del Cinvestav en Irapuato.
****Rafael Montiel
Doctor en Biología y dirige el Laboratorio de Interacción Núcleo-Mitocondrial y Paleogenómica de la Unidad de Genómica Avanzada del Cinvestav.
Fuente: Revista Avance y Perspectiva