Arnaud Ronceret
El Dr. Ronceret estudió el doctorado en genómica y desarrollo de plantas en la Universidad de Perpiñán, Francia. Hizo un posdoctorado en la Universidad Cornell, EEUU. Es Investigador en el Instituto de Biotecnología – UNAM. Apasionado por el proceso biológico fundamental de la meiosis al nivel molecular en plantas y particularmente en el maíz. Es miembro de la Academia de Ciencias de Morelos.
Esta publicación fue revisada por el comité editorial de la Academia de Ciencias de Morelos.
Todos venimos de una primera célula formada por un evento de fecundación, es decir de la fusión de dos células: un óvulo que viene de nuestra madre y un espermatozoide que viene de nuestro padre. Estas células transmiten nuestro patrimonio genómico, portado en nuestros cromosomas (estas entidades microscópicas que contienen nuestros genes) de una generación a la otra, mezclando el patrimonio genómico de nuestros padres.
Aunque tengamos los mismos padres, ¿qué hace que todos los hermanos o hermanas tengan rasgos distintos? La respuesta está en un proceso llamado meiosis – del cual se originan tanto los óvulos como los espermatozoides. Este artículo trata sobre el descubrimiento y el funcionamiento de la meiosis.
Antes de que se formen el óvulo y el espermatozoide existe una división celular muy peculiar que permite reducir a la mitad el número de cromosomas de las células que participan en la fecundación. La etapa de división del número de cromosomas se conoce como meiosis. En griego μείωσις [meíōsis] quiere decir ‘disminución’. Cuando se fusionan los dos gametos (el óvulo y el espermatozoide), la nueva célula fusionada (conocida como el cigoto) hereda el número propio de cromosomas de la especie. La mayoria de las células del cuerpo humano (células somáticas), conservan 46 cromosomas de generación en generación. En la especie humana, la división meiótica que ocurre en los testículos del padre genera espermatozoides con 23 cromosomas a partir de una célula que poseía 46 cromosomas. Igualmente, en los ovarios fetales de la madre hay células que realizan la meiosis para formar sus óvulos. La fecundación restablece los 46 cromosomas en el cigoto, que se multiplica en adelante por mitosis (células clonales) formando todas las células ‘somáticas’ de nuestro cuerpo (Figura 1).
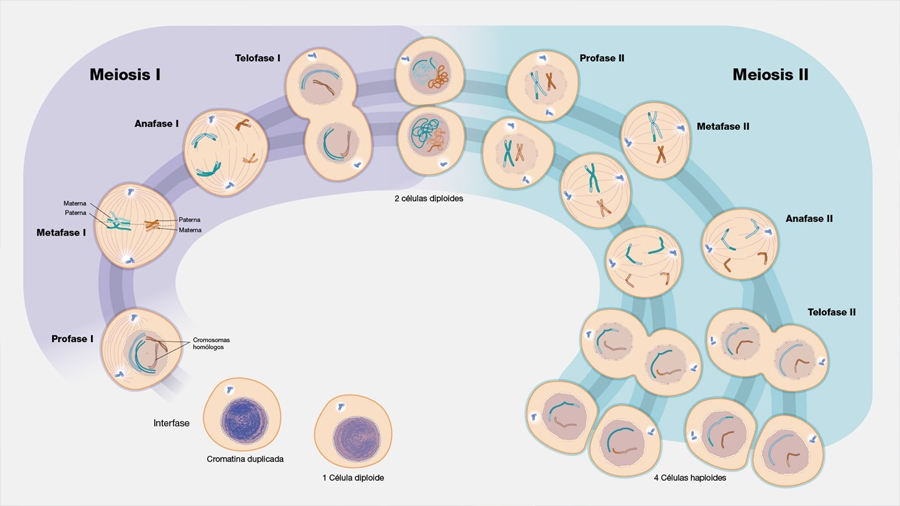
Uno podría pensar que un proceso tan central para los seres vivos que se reproducen sexualmente se descubrió con relativa facilidad. Sin embargo, esto no fue así. Lo que comprendemos actualmente se obtuvo gracias al trabajo de muchos investigadores, trabajando con diferentes organismos. Acompáñenme en un viaje histórico para descubrir cómo se alcanzó este conocimiento.
¿Cómo se descubrió este proceso?
El ciclo de vida que alterna las generaciones (Figura 2) es conocido desde 1851 gracias a la investigación del citólogo (biólogo que estudia las células) Wilhelm Hofmeister (1824-1877) que trabajó con plantas basales como musgos y helechos (Figura 3A). Hofmeister observó las estructuras celulares de varios tejidos vegetales teñidos con colorantes que permiten observar sus constituciones al microscopio óptico. De sus meticulosas observaciones, realizó una notable síntesis: todas las especies vegetales tienen alternación de generaciones. Varios biólogos consideran este descubrimiento uno de los más importantes en la biología fundamental del siglo XIX, junto con la teoría celular y la teoría de la evolución. El ciclo vital se cumple en todos los organismos con vida sexual: animales, plantas y hongos que presentan una gran diversidad de formas dentro de cada especie (Figura 2). Este ciclo vital de las especies sigue el orden cigoto, células somáticas, gametos, fecundación por fusión de dos gametos, cigoto. Sin embargo, Hofmeister no entendía cómo se transmitían y dónde residían los rasgos heredables cuando se formaba un cigoto a partir de la fusión de dos gametos y qué podía crear esta diversidad entre individuos de una misma especie.
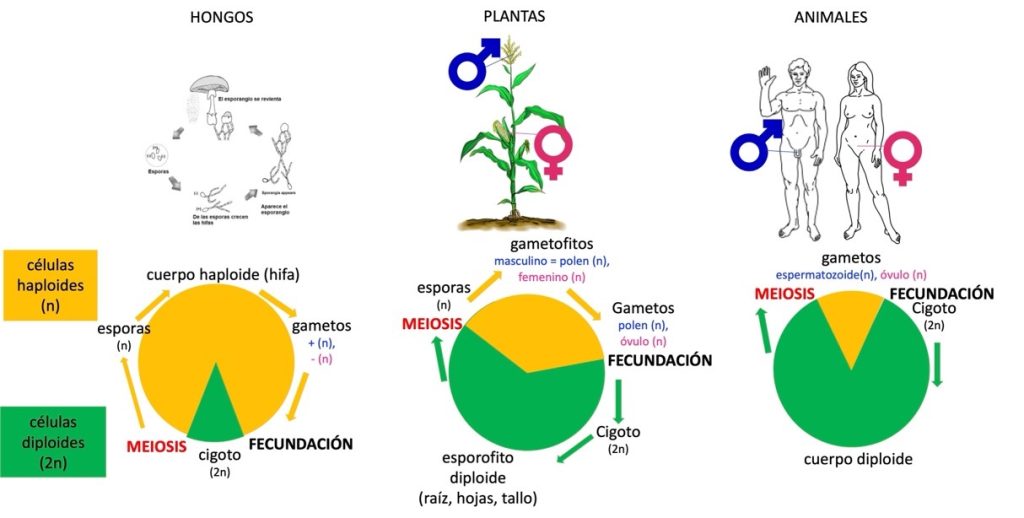
La naturaleza y proceso de formación de los gametos motivó intensas investigaciones citológicas desde el último cuarto del siglo XIX, mucho antes de saber que cada cromosoma está formado por una molécula de ADN compactada y asociada con varias proteínas.
En 1876 la meiosis fue descubierta y descrita por primera vez en los huevos del erizo del mar por Oscar Hertwig (1849-1922) (Figura 3C) y a partir del 1883, el citólogo Eduard Van Beneden (1846-1910) (Figura 3E), estudió la fecundación en Ascaris megalocephala, un nematodo parásito del intestino de los caballos, que solamente tiene dos pares de cromosomas. Gracias a esta especie, donde pueden reconocerse con facilidad los cromosomas individuales, van Beneden describió en 1888 cómo la meiosis reduce el número de cromosomas antes de formar los gametos. En 1890, August Weismann (1834-1914) (Figura 3D), interpretó teóricamente que el número de cromosomas (conocido ahora como diploide) de una célula somática tiene que partirse a la mitad durante la fase reproductiva del ciclo de vida de los organismos. La meiosis permite que se forme un germoplasma (una célula con un número de cromosomas reducido a la mitad, conocido ahora como haploide) antes de formar los gametos. En las células germinales (ahora conocidas como meiocitos) que van a formar los gametos, el número de cromosomas se reduce a la mitad mediante dos divisiones celulares sucesivas. En general, el número de cromosomas de células somáticas diploides es par, mientras el número haploide de cromosomas de los gametos puede ser impar.
Debido a varios problemas técnicos de observaciones microscópicas, la descripción de como se comportan los cromosomas en las diferentes etapas de la meiosis necesitó más de medio siglo (de 1876 a 1932) para entender claramente que las diferentes etapas meióticas corresponden a dos divisiones celulares sucesivas. Este entendimiento progresivo del proceso meiótico a través de minuciosas observaciones citológicas coincide con el re-descubrimiento en 1901 de las leyes de Gregor Mendel (1822-1884) (Figura 3B) publicadas en 1866. La disciplina que estudia la transmisión de los caracteres heredados se llama Genética. Casi concomitante, la teoría cromosómica de la herencia publicada en 1903 por Theodor Boveri (1862-1915) (Figura 3F) y Walter Sutton (1877-1916) (Figura 3G) permitió relacionar los cromosomas con el material biológico que transmite la totalidad de los rasgos individuales que se heredan (genes) entre generaciones. Los cromosomas parentales se comportan durante la meiosis como lo predice las leyes de Mendel, segregándose de manera independiente uno del otro durante la formación de los gametos, y reconstituyendo un par de cromosomas homólogos en el cigoto.

En gran parte, la dificultad para interpretar las observaciones de las divisiones meióticas en células germinales se generó porque los meiocitos han previamente duplicado su material genético, exactamente como el inicio de cualquier otra división celular (mitótica). Desde un punto de vista matemático, la división meiótica corresponde a una división por dos del número de cromosomas. Sin embargo, los mecanismos biológicos que realizan esta reducción cromosómica siguen los primeros pasos de una división mitótica, solo que se realizan después dos divisiones sucesivas en lugar de una. Así, durante la meiosis, los cromosomas son duplicados (x2) antes de ser divididos dos veces (/4). Claramente, el proceso meiótico deriva evolutivamente de un proceso mitótico. Además de esta adaptación que realiza dos divisiones sucesivas, las etapas iniciales de la meiosis (profase I) presentan también adaptaciones meióticas especificas en comparación de una división mitótica. La profase I de la meiosis es sin duda la etapa mas dinámica de la vida de una célula. Los cromosomas meióticos entran en un frenesí de movimientos para asociarse correctamente de dos en dos antes de poder separarse de manera equitativa durante la primera división meiótica. La segunda división meiótica se parece a una división mitótica: las cromátidas hermanas (que corresponden a las dos moléculas de ADN inicialmente duplicadas) se separan cada una a un polo de la célula antes de que la célula se divida. Estas dos divisiones celulares sucesivas forman cuatro células (una tétrada) que contiene cada una la mitad del número de cromosomas (23 en humanos). Estas tétradas son las células que pueden diferenciarse en gametos.
Los primeros citólogos que analizaron la meiosis estuvieron intrigados para saber cómo los cromosomas homólogos, siendo unidades microscópicas, podían asociarse con tanta precisión. En todas sus preparaciones citológicas, se observaba la especificidad de asociación entre cada cromosoma grande y cada cromosoma pequeño. Ahora sabemos que cada cromosoma paterno se asocia solamente con su homólogo materno. Durante la profase I de la meiosis todos los cromosomas homólogos se reconocen y se asocian de manera selectiva únicamente con su homólogo, para formar lo que se conoce como los bivalentes.
En 1905, Nettie Stevens (1861-1912) (Figura 3I) y Edmund Beecher Wilson (1856-1939) (Figura 3H) descubrieron los cromosomas sexuales que determinan el sexo biológico, en un insecto coleóptero del género Tenebrio y en varios insectos Hemiptera, respectivamente.
Durante la meiosis en las mujeres (las cuales contienen dos cromosomas sexuales iguales, denominados XX) el alineamiento de los dos cromosomas sexuales ocurre a lo largo de toda su longitud. En los hombres, que contienen los cromosomas sexuales X y Y, sus cromosomas sexuales forman un bivalente particular durante la meiosis, debido a que estos no son de igual longitud. El cromosoma Y es más pequeño que el cromosoma X. Sin embargo, sus asociación y recombinación en su parte común es necesaria para formar el bivalente sexual y la segregación equilibrada en las tétradas. La segregación de un cromosoma X o Y en los espermatozoides humanos es lo que determina el sexo en su descendencia. En la especie humana, los óvulos no determinan el sexo de los bebés (solo pueden llevar un cromosoma X), mientras que los espermatozoides pueden contener un cromosoma X o uno Y. Así, los espermatozoides que tienen un cromosoma Y, al encontrarse con un óvulo conteniendo el cromosoma X, determinan que se formará un cigoto XY, produciendo un hombre. Si el espermatozoide contiene un cromosoma X, el cigoto resultante será XX, produciendo así una mujer. Esta segregación aleatoria de los cromosomas X y Y en los espermatozoides es el mecanismo biológico que asegura que la cantidad de hombres sea igual al número de mujeres en la población humana.
Al estudiar los enormes cromosomas meióticos de varias especies de tritones y salamandras, claramente visibles bajo microscopía óptica, François Alfons Janssens (1863-1924) (Figura 3J) descubrió que, además de unirse, los bivalentes intercambian porciones de brazos cromosómicos en sitios con forma de X (letra griega ‘Ji’) a los que llamó quiasma (quiasmata en plural). Sus observaciones e interpretaciones correctas de los intercambios de porciones de cromosomas homólogos durante la meiosis fueron publicados como la ‘teoría de la chiasmatipie’ en 1909 (Figura 4).
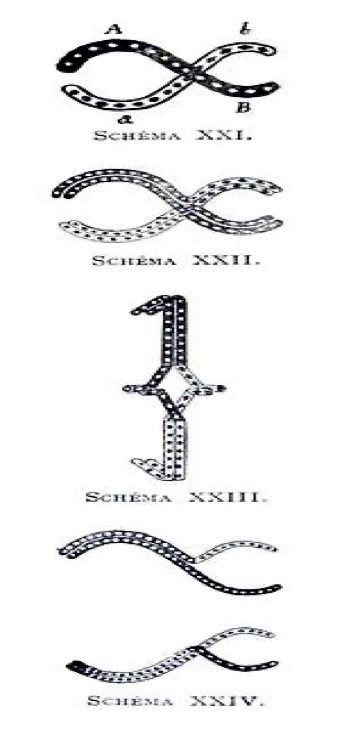
Gracias al citólogo Edmund B. Wilson (que conocía la publicación en francés de Janssens), Thomas Hunt Morgan (1866-1945) (Figura 3K), su vecino trabajando en la famosa Fly room (habitación de las moscas) de la Universidad Columbia de Nueva York, interpretó correctamente sus datos genéticos usando la mosca de la fruta Drosophila melanogaster (literalmente ‘amante del rocío de vientre negro’) desde 1916. Thomas Morgan, y su esposa Lilian (1870-1952) (Figura 3L), habían identificado en 1910 el primer mutante espontáneo que afecta un gen (white o w) cambiando el color de los ojos de esta mosca de rojo a blanco, pero solamente en los machos. La Drosophila, que puede producir docenas de generaciones en un solo año, fue el modelo ideal para los primeros genetistas. Este modelo permitió de entender las leyes de transmisión de los genes y correlacionarlos con la transmisión de los cromosomas, porque Drosophila tiene solamente cuatro pares de cromosomas incluyendo un cromosoma sexual. La observación que solo los mutantes w machos tenían ojos blancos permitía de posicionar el gen white en el cromosoma sexual X, y descubrir la herencia ligada al sexo. Los estudiantes de Thomas H. Morgan, Calvin Bridges (1889-1938) (Figura 3M), Herman Müller (1890-1967) (Figura 3N) y Alfred Sturtevant (1891-1970) (Figura 3O) descubrieron rápidamente cómo los genes son alineados en los cromosomas como ‘perlas en un collar’ y cómo los cromosomas de dos padres pueden intercambiar sus genes y formar nuevas combinaciones a partir de sitios de entrecruzamientos genéticos. Si los cromosomas de dos padres se comparan a collares de perlas de diferentes colores, como blanco y negro, existe descendientes que heredan collares compuestos de grupos de perlas negras y grupos de perlas blancas (Figura 5) para crear una nueva combinación entre cromosomas homólogos parentales (ahora conocida como recombinación intra-cromosómica). Dependiendo en donde se hace los puntos de entrecruzamientos (o ‘crossing-over´ en inglés), la composición de los cromosomas formados durante la meiosis corresponde a un mosaico especial de distintos genes transmitidos entre cada hermano.
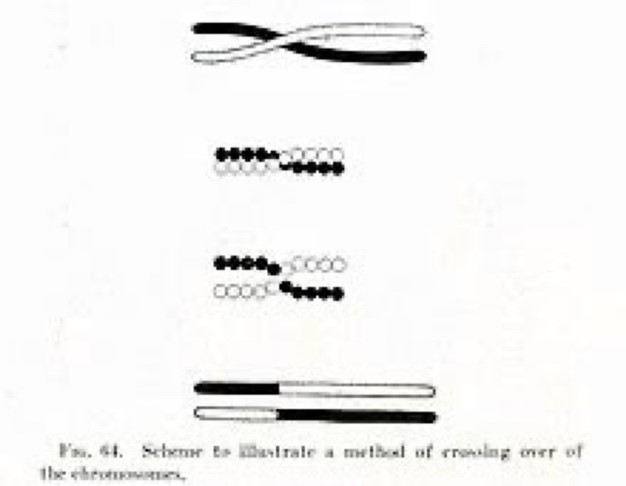
Usando una colección de varios mutantes obtenidos en Drosophila, Stutervant tiene la brillante idea de usar la frecuencia de recombinación entre mutantes para estimar la distancia entre los genes correspondientes y mapear cómo se posicionan los genes uno al otro. La unidad de distancia genética (el porcentaje de recombinantes) se le denomina centi Morgan (cM) en homenaje a su tutor. Un cM corresponde a 1% de descendientes recombinantes. Así desde 1913 se generan los primeros mapas genéticos en Drosophila. Herman Müller se da cuenta que dos eventos de entrecruzamientos no son independientes y que la aparición de un entrecruzamiento en una región del cromosoma da menos probabilidad de formar otro entrecruzamiento en su vecindad. Müller llama este importante fenómeno de la recombinación meiótica como interferencia entre entrecruzamientos en 1916.
Estas observaciones genéticas se revelan como válidas entre diferentes animales, plantas y hongos en las décadas siguientes y son generalizadas y validadas hasta en humanos. En 1926 Morgan y sus estudiantes publican el libro ‘La teoría del gen’ que explica cómo los genes son alineados en los cromosomas y cómo nuevas combinaciones genéticas y cromosómicas se forman durante los procesos de meiosis y fecundación, explicando la diversidad y la unicidad de los individuos que se reproducen de manera sexual. El papel de la sexualidad celular fue claramente establecido desde 1926: los genes son portados en los cromosomas como perlas en un collar y se pueden recombinar como lo había notado Janssens desde 1909 a nivel citológico. Así el proceso meiótico que genera gametos puede crear la unicidad entre hermanos, es también la fuente de la enorme diversidad observada entre especies que usan la sexualidad como modo de reproducción.
En dos próximos artículos de La Ciencia, desde Morelos para el mundo, explicaré los avances esenciales que se realizaron entre 1926 – 1983 y desde 1983 hasta la actualidad. Explicaré sobre los mecanismos que permiten la formación de estos entrecruzamientos a nivel molecular y del ADN y qué implicaciones médicas tienen estos conocimientos.
Para saber más
- Origen de las fotos:Todas las fotos vienen de Wikipedia, excepto las de van Beneden que viene de la Universidad de Lieja (https://200.ulg.ac.be/vanbeneden.html, la de François Alfons Janssens que viene de https://en.geneanet.org/media/public/frans-janssens-1863-1924-2-27977321, la de Calvin Bridge que viene de Getty Images https://www.gettyimages.ca/detail/news-photo/dr-calvin-bridges-of-the-carnegie-institute-carrying-on-news-photo/515132994, la de Thomas Hunt Morgan (https://www.nobelprize.org/prizes/medicine/1933/morgan/article/) y de Herman Muller (https://www.nobelprize.org/prizes/medicine/1946/muller/biographical/ ) que viene del sitio del Premio Nobel. La foto de Alfred Stutervant viene del sitio de Cold Spring Harbor https://dnalc.cshl.edu/view/16282-Gallery-11-Alfred-Sturtevant-1918.html.
- Portal académico CCH-UNAM. Meiosis. https://e1.portalacademico.cch.unam.mx/alumno/biologia1/unidad2/meiosis/meiosis
- CUAED/Bachillerato a Distancia – UNAM. Meiosis. https://uapa.cuaed.unam.mx/node/616
- UAPA, Facultad de Medicina, UNAM. Meiosis. https://repositorio-uapa.cuaieed.unam.mx/repositorio/moodle/pluginfile.php/2489/mod_resource/content/6/UAPA-Meiosis/index.html
Fuente: acmor.org